Кровеносная система
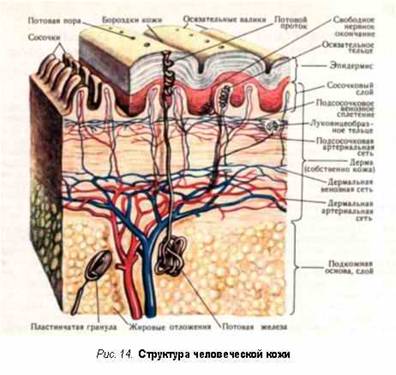 Возьмем
еще кровеносную систему. От сердца идут длинные ветви сосудов. Общая длина их в
одном человеке почти втрое превышает периметр земного шара. Артерии уносят от
сердца свежую, обогащенную кровь, вены приносят обедненную. Ни один насос не
способен прокачивать ежесекундно 60‑90 мл жидкости через настолько развитую
сетку сосудов. - А этого и не требуется из-за особого строения системы: сердце
создает небольшую разность давления между артериями и венами, а капилляры,
располагающиеся между ними, благодаря капиллярному эффекту (диаметр которых
несколько микрометров), сами берут необходимое количество артериальной крови и
сами передают ее к венам. Капиллярные стенки прозрачны, так что каждая клеточка
всегда имеет неограниченный резервуар питательных веществ. Проложить к
каждому участку тела свой акведук - неужели это работа мутаций? А ведь нужно
еще регулировать сосудистое давление!.. (Наиболее ярко важность регулировки
давления видна в случае жирафа: если бы не мгновенное срабатывание системы
клапанов, кровь просто разорвала бы жирафу голову при резком опускании головы.)
Возьмем
еще кровеносную систему. От сердца идут длинные ветви сосудов. Общая длина их в
одном человеке почти втрое превышает периметр земного шара. Артерии уносят от
сердца свежую, обогащенную кровь, вены приносят обедненную. Ни один насос не
способен прокачивать ежесекундно 60‑90 мл жидкости через настолько развитую
сетку сосудов. - А этого и не требуется из-за особого строения системы: сердце
создает небольшую разность давления между артериями и венами, а капилляры,
располагающиеся между ними, благодаря капиллярному эффекту (диаметр которых
несколько микрометров), сами берут необходимое количество артериальной крови и
сами передают ее к венам. Капиллярные стенки прозрачны, так что каждая клеточка
всегда имеет неограниченный резервуар питательных веществ. Проложить к
каждому участку тела свой акведук - неужели это работа мутаций? А ведь нужно
еще регулировать сосудистое давление!.. (Наиболее ярко важность регулировки
давления видна в случае жирафа: если бы не мгновенное срабатывание системы
клапанов, кровь просто разорвала бы жирафу голову при резком опускании головы.)
Нервная система
А такая вещь, как нервная система,
мозг - неужели это тоже продукт мутаций? Тогда значит и сознание - продукт
мутаций?
Еще Зенон Элейский (490-430 до н.э.)
и Демокрит (460-371 до н.э.) принципиально разграничивали чувственное и
рассудочное (аналитическое). Все же, вопреки имеющейся принципиальной разнице,
природа их сходна, поскольку аналитическое в глубине напоминает своего рода
чувственное. Сознание базируется, с одной стороны, на системах чувствования и,
как их следствие, эмоциях. С другой стороны, оно опирается на внутреннее
"видение" (анализ, синтез, наблюдение, абстрагирование,
распознавание категорий, идентификация отношений и т.п.), которое
также можно назвать своеобразным чувством: мы ощущаем, что один предмет
больше другого, что одно качество/мера выше другого и т.п.
Умственное и чувственное тесно
переплетены в сознании. Даже интуиция - разве это тоже не своеобразное чувство,
не распознавание/чувствование наиболее глубинных принципов и (мета)образов?
Сознание является единым целым, оно есть приемником, настроенным на
детектирование/распознавание всех моментов действительности, на распознавание
действительности как целого. (Единое целое, настроенное на распознавание единого
целого.) Одни участки сознания распознают более грубые моменты ("знаки")
действительности, другие - более тонкие. Причем, высшие каскады принимают на
вход "первично-обработанные знаки" со выхода предыдущих (низших) каскадов,
вырабатывая более сложные "(мета)знаки", так что имеется распределение
образов/знаков на "внешние" и "внутренние" (по источнику происхождения).
Чувствование примитивных "образов", как, например, теплое/холодное, и
"чувствование" высочайших абстракций, как, например,
справедливое/несправедливое, принципиально подобны. Разделение единого целого
на чувственное, рассудочное и интуитивное весьма условно.
Разумеется, "аналитическое чувство"
не является обычным чувством. При "видении" сознание оперирует образами и
ситуациями [Ситуация
- совокупность образов и связей/отношений
между ними], - и решение
вырабатывается тоже в виде образа или ситуации. Если бит является
единицей "компьютерной/технической" информации, то единицей "природной"
информации является элементарный образ/знак, минимальная единица "ситуационной
информативности".
Аналитическая часть мозга и компьютер
во многом подобны между собой. Различие между ними состоит в кардинальном
отличии основополагающих механизмов и алгоритмов. Мозг обрабатывает "за один
такт" несколько образов, целую ситуацию, а компьютер - всего несколько бит, пару
чисел [Проф. И.И. Клетченков, частный
разговор (1995)].
С компьютером все значительно примитивнее, чем могло казаться: элементная база
арифметико-логического блока микропроцессора может состоять всего из двух
функциональных элементов "и-нет" /
"или-нет", а все операции над
числами - сведены к операциям логического сложения, логического вычитания и
инверсии. В случае же мозга, аналитические процессы так тонко спрятаны и
настолько далеки от всего известного, что даже подступиться к себе не дают.
Как происходит оперирование образами и как "само собой" вырабатывается решение
ситуаций - это большая тайна, сверхсложный алгоритм.
Психика и сознание - целая отдельная
Вселенная с психическими состояниями, категориями, понятиями, желаниями, целями
и идеями. Что такое Я? Кто когда-либо видел собственное Я? Как его
себе вообразить? Трудность для нашего сознания вопроса о "форме" и
"местонахождении" самого себя является огромным парадоксом, свидетельствующим о
чрезвычайной сложности устройства. И очевидно, - неслучайного устройства. А что
такое сон? Существенного прогресса в подобных размышлениях не видно. -
Так сложны мозговые процессы!
Психологи с нейрофизиологами топчутся
на месте в понимании "механизма чувствования", равно как физики - в понимании
"структуры поля". (Можно лишь посочувствовать, ведь это наифундаментальнейшие
вещи в соответствующих областях.) - Настолько сложным есть устройство
человеческого интеллекта!
То, что животные имеют меньший мозг,
не означает слабость их мозга. Просто у каждого своя специализация. Щука
-высококлассный специалист по карасям, а зайцы - мастера экстра класса по бегу
на пересеченной местности. Паук (можно ли разглядеть его крохотный мозг?) очень
хорошо умеет плести паутину и дежурить на ней. А для описания
коллективно-координированных умений пчел и муравьев просто не хватает тут места.
Полноценного понятийного мышления животные не имеют, но они способны ко всем
другим типам мышления:
"И чувственно-образное, и техническое
мышление, очевидно имеются и у высших животных (обезьян, собак, котов, дельфинов
и т.п.). Доказано, что высшие животные пользуются всеми известными обычной (то
есть формальной) логике методами - дедукцией, индукцией, синтезом, анализом,
экспериментом, абстрагированием и т.п. - вплоть до образования родовых понятий,
то есть всеми теми методами, которыми используется человек." [Енгельс
Ф. ДЁалектика природи. - К., 1977.
- С. 201-202, как
перефразировано в: Кочерган М.П. Загальне мовознавство. - К.: АкадемЁя,
1999. - С. 39-40]
Все животные рождаются чему-то
"наученными" или готовыми к успешному овладению определенной "наукой". Они все
много чего знают и умеют. Но - только то, что им предназначено. Обезьяна,
к примеру, никогда не заговорит, поскольку
ей не дано соответствующего
речевого участка мозга. Она может понять внешнее слово (иногда даже
предложение), но сама не сможет составить предложение - ей недосягаемы понятия
синтаксис и грамматика. То же и с "речью" попугая.
Неужели сложность мозга есть
продуктом мутаций? А
кровеносной и нервной системами далеко не исчерпывается перечень систем
организма. И все они работают очень слаженно: та же кровеносная система тесно
сотрудничает с дыхательной, эндокринной, лимфатической, пищеварительной и,
конечно, нервной системами.
Примечание:
Лимфатические сосуды собирают "лишнюю" кровяную
сыворотку и пульсациями направляют ее к венам. У многих животных есть
специальные лимфатические сердца.
Попытки согласовать дарвинизм с
данными палеонтологии
Теория Дарвина является теорией
постепенности, градуализмом. Градуализм постулирует накопление многих
сравнительно маленьких телесных изменений таким образом, что в результате
получается новая полезная структура (орган или система). Градуализм - это
гипотеза постепенности развития новых структур небольшими шагами.
Тем не менее, учитывая чрезвычайную
ограниченность коллекции вымерших видов, которые можно было бы рассматривать как
промежуточные межвидовые звенья, многим палеонтологам 1950х-60х
годов стало ясно, что градуализм исчерпал себя как не подтверждающийся фактами.
Для теории Дарвина настал критический момент: осуществился наихудший сценарий
относительно подтверждения гипотезы постепенного развития видов из единого
предка - не было выявлено ни одной убедительной промежуточной формы.
Ответом материалистов на вопрос о
механизме видовой эволюции стала концепция пунктуализма, согласно которой
образование новых видов происходит не на протяжении многих миллионов лет, а в
значительно более короткие сроки - от тысячи до нескольких десятков тысяч лет,
что выглядит лишь точкой/пунктом в сравнении с десятками миллионов лет.
Американские палеонтологи С. Гоуд и Н. Елдридж предложили эту концепцию в 1972
году и назвали ее теорией скачкообразного равновесия. За скачкообразным
формированием новой полезной структуры, а значит, и нового вида, следует
сравнительно длинный стационарный период. Далее прыжком образуется новый вид, за
ним снова продолжительное равновесие, и так далее. Эволюция видов в этой
гипотезе больше напоминает движение вверх ступеньками, чем движение вверх
вдоль наклоненной плоскости. Тем, что продолжительность прыжка мала по сравнению
с продолжительностью стационарного состояния, можно объяснить отсутствие среди
окаменелостей четких филогенетических линий: Малочисленные промежуточные формы,
подобно хрупкому мостику от вида к виду, "упали" в пропасть времени и там
безнадежно затерялись.
Авторы этой подкорректированной
теории Дарвина предполагают, что при изоляции определенной периферийной
популяции, в ней может спонтанно состояться нарушение равновесия, например,
вследствие существенного изменения внешних условий, и этого нарушения достаточно
для сравнительно быстрой "адаптации" организмов к новым условиям. Источником
"адаптации", то есть движущей силой развития новых полезных структур, видятся
мутации.
Однако, как было показан выше,
механизм мутаций не является действенным ни в сжатые сроки, ни на протяжении
миллионов лет, поскольку не содержит объяснения закрепления промежуточных
мутационных этапов. Мутации же без промежуточных этапов, при которых за одну
мутацию появляется, например, целая система слуха или нюха, то есть
макромутации, - более чем сомнительны. Покажем это следующими расчетами.
Вероятность удачной перестановки
элементов ДНК "макромутационного" масштаба сопоставима с вероятностью новой
самосборки ДНК (10‑40000, см. раздел 1.2). Пусть даже она будет в
10000000000 раз более благоприятной/большей, то есть 10‑39990. Для
оценки реалистичности появления конкретной комбинации надо определить частоту
появления новых комбинаций. Как уже упоминалось, темп наследственной мутации
(появление наследственной "муто-комбинации" ДНК) составляет меньше единицы на
миллион новорожденных. Примем, что в популяции каждый день рождается миллион
особей, - тогда частота появления мутантов в день будет меньше единицы (10‑6
· 106 = 1). Значит, для появления одной удачной макромутации нужно 1039990
дней или 1039987,4 лет (для сравнения, сто миллиардов лет
записывается цифрой 1011). Появление хотя бы одной новой
полезной структуры в одной популяции за 100 миллиардов лет является событием
математически невероятным (10‑39976,4). А за 10 тыс. лет,
требуемых пунктуализмом, - еще более невероятным (10‑39983,4).
С одной стороны, для микромутаций
нет механизма их закрепления, а с другой, макромутации являются событиями
безмерно маловероятными. Ни микро-, ни макромутации не производят новых полезных
структур.
Таким образом, механизм мутаций не
обеспечивает теоретического объяснения возникновения полезных структур, не
является логичным на фоне уровня сложности кода ДНК, и его действенность не
подтверждается мутационными экспериментами.
Механизм мутаций, таким образом, не действенен.
Естественный отбор
Кроме мутаций и естественного отбора,
в "формуле эволюции" фигурируют еще такие факторы, как половое
воспроизведение, генетический дрейф, географическая изоляция
и, в случае человека, социальное регулирование брака [Dobzhansky
T.
Genetics and the
Origin of Species
- N.Y.:
Columbia
Univ.
Press,
1951. - P.
304].
Но весом этих факторов можно пренебречь в сравнении с двумя главными (мутации и
естественный отбор).
По эволюционной схеме, естественный
отбор вступает в действие после механизма мутаций. Он не действенен сам
по себе - это признал в 6ом издании сам Дарвин. Естественный
отбор должен подхватывать "удачно" трансформированный ген и выводить его в
лидеры, все увеличивая число организмов, в которых он присутствует -
организмов-мутантов. Два фактора, считают неодарвинисты, работают не в
отдельности, а лишь в тандеме. Каждый из факторов в отдельности бессилен.
Поскольку механизм мутаций не
действенен, то и естественный отбор не действенен:
ему нет, чего подхватывать. На этом тезисе рассмотрение естественного отбора как
столпа теории эволюции видов можно закончить.
Поговорим о сущности естественного
отбора и смешении двух разных понятий под словом эволюция. Но сначала о
ситуации с терминами.
С понятием, стоящим за термином
вид, в биологии не все так просто. Таксономия и систематика, занимающиеся
классификацией организмов, не имеют четких объективных критериев размежевания
видов. Часто видом называют популяцию организмов, способных к полному
скрещиванию (с рождением небесплодных потомков).
Бык и буйвол - это два отдельных вида
или один? То же самое - собака и волк? Биологи считают их отдельными видами, а
тем не менее, они могут скрещиваться. Виды объединяются в роды, а роды - в
семьи. Есть некоторая неясность в том, что относить к роду, а что к виду.
Дополнительно затуманивает вопрос явление потери способности скрещиваться
популяциями одного вида при продолжительном географическом разъединении:
например, дятлы Европы и Урала или лягушки разных регионов. Внешне вроде бы одна
единица, но плохо скрещиваются, - и уже поэтому могут считаться разными видами.
А может, при рассмотрении вопроса
происхождения видов лучшее пользоваться другим термином? Полвека тому назад было
введено понятие базовый тип, под которым понимаются все организмы,
способные между собой к "наименьшему" скрещиванию (то есть, если оплодотворенная
яйцеклетка "держится" хотя бы несколько делений). Кое-кто предлагает
пользоваться термином барамин, исходя из креационной модели, согласно
которой, все живое было создано "по роду своему". Неизвестно, насколько барамин
тождествен базовому типу. Например, лев и кот являются одним базовым типом, но
есть сомнение, что они являются одним барамином. Аналогично, всех ли лягушек и
всех змей считать одной категорией? В этом есть вопрос.
Одно ясно - данные палеонтологии
решительно свидетельствуют об отделенности каждого базового типа организмов.
Между базовыми типами нет промежуточных форм, то есть преобразование лягушек в
змей или рептилий в птиц невозможно.
Вместо единого дерева видов мы имеем
совокупность отдельных, независимых деревьев-видов - видовой лес. Каждому
отдельному дереву в этом лесу отвечает один барамин или базовый тип. Например,
можно представлять себе дерево псовых, ветвями которого являются собаки, волки,
динго и койоты (возможно, также шакалы и лисы); или дерево слоновых, ветвями
которого являются африканские слоны, индийские слоны, мамонты и мастодонты; или
дерево рогатого скота с ветвями - быки, буйволы, зубры, брахманы, яки, гаури,
гаяли и бизоны. К дереву кошачьих можно попробовать зачислить домашних и диких
котов и, возможно, рысей и пум. Большие кошачьи (львы, тигры, леопарды)
представляют собой либо отдельное дерево, либо раннюю ветвь дерева кошачьих [Если
барамин кошачьих настолько широк (что очень сомнительно)].
К дереву медведевых, наверное, относятся черные, серые (гризли), бурые,
полярные, пещерные медведи и, возможно, панды. Жирафы, верблюды и страусы
являются безветочными деревьями жирафовых, верблюдовых и страусовых,
соответственно.
Можно предположить, что базовый тип
или барамин способен порождать новые виды: в рамках дерева рогатого скота,
наверное, возможно образование "быко-зубра", а в рамках дерева оленевых -
"лосе-оленя". Порождение же новых базовых типов, то есть новых деревьев,
невозможно. Деревья лишь исчезают, видовой лес редеет, а не наоборот.
Естественный отбор не приводит к
усложнению вида, так как мутации недейственны, - но он не разрешает и
деградировать виду, так как слабейшие гибнут. Сущность естественного отбора - в
консервации. Это механизм консервации здорового в виде.
Изменчивость пород/разновидностей
эволюционисты называют микроэволюцией. Она состоит в диверсификации
пород/разновидностей в рамках одного вида (или видов в рамках рода) под влиянием
естественного или искусственного отбора. Микроэволюция, в отличие от
макроэволюции (собственно эволюция или неодарвинизм), действительно, имеет
место. Пример микроэволюции: все то разнообразие перьев и клювов в видах
вьюрковых, которое наблюдал Дарвин на Эквадоре, можно получить из одной
единственной популяции путем искусственного отбора за десятки лет, - в
противоположность десяткам тысяч лет, необходимым естественному отбору.
Другое дело - макроэволюция, для
которой требуется не изменение комбинации доминантных/рецессивных позиций генов
в заданном наборе, а образование нового генетического набора (генофонда).
Макроэволюция оказывается невозможной:
"Если под эволюцией мы понимаем
макроэволюцию (что мы в дальнейшем и будем делать), то наикатегоричнейшим
образом можно сказать, что [эволюционное] учение полностью лишено научной
санкции. ...По сей день не существует ни малейшего бона фиде ["достойный
доверия", лат.] научного доказательства в поддержку тезиса, будто
макроэволюционные трансформации когда-либо имели место."
[Известный
математик и физик Дж.
Смит
- Smith J.W.
Teilhardism and the New Religion: A Thorough Analysis...
- Rockford
,
Illinois
,
1988.
-
Р. 5-6]
Съезд
ведущих эволюционистов мира обсуждал вопрос
" могут ли механизмы, лежащие в основе
микроэволюции, быть экстраполированы для объяснения явления макроэволюции."
- Ответом было "категорическое нет" [Lewin
R.
Evolutionary Theory
Under Fire //
Science
- Vol.
210, No.
4472 (
Nov.
21,1980
) -
Р.
883].
"Микроэволюция
- да. Макроэволюция - нет! Это природный факт (то есть, это можно
продемонстрировать) огромной важности..."
[Известный
биолог
проф.
Ф.
Марш
- Marsh F.L.
Genetic Variation, Limitless or Limited? // Creat.
Research Soc.
Quart.
- No.
4 (1983) - P.
206]
Важно понять, что микроэволюция,
по сути, не является эволюцией. Микроэволюция означает гибель
малоприспособленных и выживание лучших представителей популяции без образования
новых органов. То есть, это "0" или "–". Эволюция же требует образования нового,
новых органов, это "+". Таким образом, эволюции нет ни в макроэволюции, ни в
микроэволюции: эволюции нет вообще, поскольку новые полезные структуры не
образуются никогда.








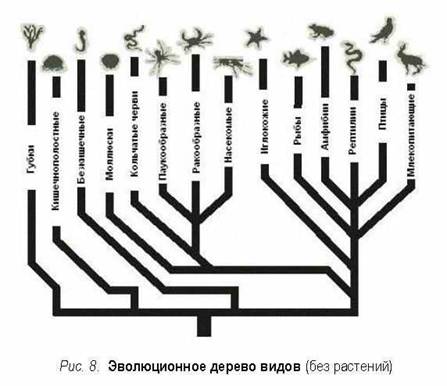 Дерево видов является попыткой
исторической реконструкции: современные виды отвечают
наиболее внешним ветвям дерева, тогда как все внутреннее есть давними видами,
гипотетической историей. Двигаясь извне к центру кроны дерева, ветви
все более сливаются, упираясь наконец в единый ветвь-ствол. Этим первоначальным
видом-стволом (общий для всех видов предок) было амебоподобное существо,
которое, возможно, уже не существует. От ствола расходятся несколько главных
ветвей, две известнейшие из которых называются животными и растениями,
а остаток рассматривается как "растения-животные".
Дерево видов является попыткой
исторической реконструкции: современные виды отвечают
наиболее внешним ветвям дерева, тогда как все внутреннее есть давними видами,
гипотетической историей. Двигаясь извне к центру кроны дерева, ветви
все более сливаются, упираясь наконец в единый ветвь-ствол. Этим первоначальным
видом-стволом (общий для всех видов предок) было амебоподобное существо,
которое, возможно, уже не существует. От ствола расходятся несколько главных
ветвей, две известнейшие из которых называются животными и растениями,
а остаток рассматривается как "растения-животные". 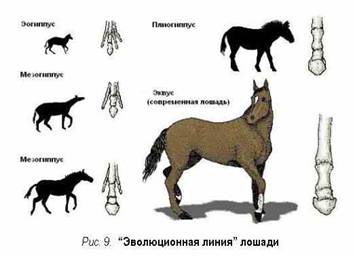 "Униформная, постепенная трансформация
хиракотериума [эогиппус,
первобытная лошадь] в эквус
[современная лошадь], столь дорогая сердцам авторов учебников, никогда не
имела места в природе. ... Эволюция семьи конеподобных, эквиде, сегодня
известна не лучше многочисленных других групп организмов..." [Палеонтолог и ярый
эволюционист Дж. Симпсон -
Simpson G.G.
Life of the
Past. -
New Haven
,
Connecticut
, 1953. -
P.
125,
127 (подчерк.
добавлено)]
"Униформная, постепенная трансформация
хиракотериума [эогиппус,
первобытная лошадь] в эквус
[современная лошадь], столь дорогая сердцам авторов учебников, никогда не
имела места в природе. ... Эволюция семьи конеподобных, эквиде, сегодня
известна не лучше многочисленных других групп организмов..." [Палеонтолог и ярый
эволюционист Дж. Симпсон -
Simpson G.G.
Life of the
Past. -
New Haven
,
Connecticut
, 1953. -
P.
125,
127 (подчерк.
добавлено)]  О
гомо габилисе широко писалось как о звене между австралопитеком (обезьяной) и
гомо эректусом (человеком), то есть как звене, связывающем наконец обезьян и
людей. Сейчас слово-классификатор гомо считается неправильно
употребляемым к данному виду животных.
О
гомо габилисе широко писалось как о звене между австралопитеком (обезьяной) и
гомо эректусом (человеком), то есть как звене, связывающем наконец обезьян и
людей. Сейчас слово-классификатор гомо считается неправильно
употребляемым к данному виду животных.  В
реконструировании гипотетических "недостающих звеньев" большую роль играет
художественная фантазия.
В
реконструировании гипотетических "недостающих звеньев" большую роль играет
художественная фантазия.  Механизм
мутаций, согласно утверждениям неодарвинистов, является первым фактором эволюции
видов. Он может обеспечивать появление определенного преимущества мутировавшего
организма перед остальными особями. Такое преимущество выражается, например, в
виде нового полезного органа или внутренней системы или в улучшении каких-то
характеристик. Его наличие служит предпосылкой дальнейшего закрепления мутации:
"удачно" мутовавшее животное имеет больше шансов прожить дольше других, породить
больше детей и расширить свое "мутагенное" присутствие в генофонде вида/рода. В
мутациях - все для неодарвинизма: если этот механизм не действенен, тогда
исчезает источник появления новых характеристик, и естественному отбору "не
будет, чего отбирать".
Механизм
мутаций, согласно утверждениям неодарвинистов, является первым фактором эволюции
видов. Он может обеспечивать появление определенного преимущества мутировавшего
организма перед остальными особями. Такое преимущество выражается, например, в
виде нового полезного органа или внутренней системы или в улучшении каких-то
характеристик. Его наличие служит предпосылкой дальнейшего закрепления мутации:
"удачно" мутовавшее животное имеет больше шансов прожить дольше других, породить
больше детей и расширить свое "мутагенное" присутствие в генофонде вида/рода. В
мутациях - все для неодарвинизма: если этот механизм не действенен, тогда
исчезает источник появления новых характеристик, и естественному отбору "не
будет, чего отбирать".  Морфология
животных в большой степени симметрична. Прежде всего, это касается внешнего
уровня, значительно меньше - внутреннего. Возьмем человеческую руку:
симметрично от правой руки располагается левая рука, - практически полностью
идентичная. Точная симметричность. Но это внешний уровень. На внутреннем уровне
симметрия исчезает: структура кровеносных сосудов, рисунок ладони и т.п., у
каждой руки уже свои. Симметрия исчезает уже даже на внешнем уровне каждой из
рук: большой палец - а напротив мизинец; кисть - а напротив предплечье;
ноготь - а на обороте кожа. Касательно поразительной внешней симметрии,
возникает вопрос к мутациям: если бы серия мутаций и была в состоянии вытворить
одну руку, то другую руку она не могла вытворить точно напротив первой - мутация
ведь действует "наугад". Вторая рука оказалась бы размещенной немного косо, на 3
см ниже или на 2 см "заднéе": функциональность руки была бы при этом вполне
удовлетворительной, преимущество двурукого существа перед однорукими было бы
несомненным, то есть новый орган "прижился бы". Кроме того, почему на второй
руке должно появиться именно 5 пальцев, а не 6? - Что, у шестипалой кисти хуже
захват? И почему на правом предплечье именно рука, а не длинный хвост? - Ведь
давним людям как скотоводам так часто нужен был кнут под рукой - вот и был бы
хвост-кнут-аркан-хобот "всегда под рукой"! А почему нос расположен по центру
лица? - Что, при нецентральном расположении хуже ощущались бы запахи? И почему
для носа не сотворилась пара (как в случае глаз, щек или ноздрей)? - Никто же не
против "стереонюханья". А может кто-то думает, что мутация не слепая, т.е.
умная?
Морфология
животных в большой степени симметрична. Прежде всего, это касается внешнего
уровня, значительно меньше - внутреннего. Возьмем человеческую руку:
симметрично от правой руки располагается левая рука, - практически полностью
идентичная. Точная симметричность. Но это внешний уровень. На внутреннем уровне
симметрия исчезает: структура кровеносных сосудов, рисунок ладони и т.п., у
каждой руки уже свои. Симметрия исчезает уже даже на внешнем уровне каждой из
рук: большой палец - а напротив мизинец; кисть - а напротив предплечье;
ноготь - а на обороте кожа. Касательно поразительной внешней симметрии,
возникает вопрос к мутациям: если бы серия мутаций и была в состоянии вытворить
одну руку, то другую руку она не могла вытворить точно напротив первой - мутация
ведь действует "наугад". Вторая рука оказалась бы размещенной немного косо, на 3
см ниже или на 2 см "заднéе": функциональность руки была бы при этом вполне
удовлетворительной, преимущество двурукого существа перед однорукими было бы
несомненным, то есть новый орган "прижился бы". Кроме того, почему на второй
руке должно появиться именно 5 пальцев, а не 6? - Что, у шестипалой кисти хуже
захват? И почему на правом предплечье именно рука, а не длинный хвост? - Ведь
давним людям как скотоводам так часто нужен был кнут под рукой - вот и был бы
хвост-кнут-аркан-хобот "всегда под рукой"! А почему нос расположен по центру
лица? - Что, при нецентральном расположении хуже ощущались бы запахи? И почему
для носа не сотворилась пара (как в случае глаз, щек или ноздрей)? - Никто же не
против "стереонюханья". А может кто-то думает, что мутация не слепая, т.е.
умная?  До 1950х
годов считалось, что только белок имеет достаточно сложную структуру, чтобы
служить носителем информации. Поэтому, после теоретического открытия генов
ученые полагали, что участок клетки, ответственный за наследственную информацию,
имеет белковую природу. Однако, канадец Аври в 1944 году установил, что эту роль
выполняет дезоксирибонуклеиновая кислота (ДНК).
До 1950х
годов считалось, что только белок имеет достаточно сложную структуру, чтобы
служить носителем информации. Поэтому, после теоретического открытия генов
ученые полагали, что участок клетки, ответственный за наследственную информацию,
имеет белковую природу. Однако, канадец Аври в 1944 году установил, что эту роль
выполняет дезоксирибонуклеиновая кислота (ДНК).