|
 |
 |
 |
Александр Кобринский Когда начинают летать динозавры Чтобы понять каким конкретным образом эволюционирует живой мир, произведем дифференциацию всей живой биологической массы по бросающимся в глаза способа контактирования с внешней средой (с реальным объектом). Таких способа два - вегетационный и сенсорный. К вегетационному способу контактирования принадлежит растительный мир, к сенсорному (чувственному) - животный. Сенсорный контакт животного мира с внешней средой может быть или чисто инстинктивным или инстинктивно разумным. Палеонтологические данные убеждают нас в том, что эволюционирует и растительный мир и животный. Не вдаваясь в подробности этих данных и не соотнося их с существующей научной дифференциацией геологического времени, но обобщая эти подробности феноменологически, вернемся к нашей работе "Мы - мутанты" [3, с. 147-153] и рассмотрим следующую схему:
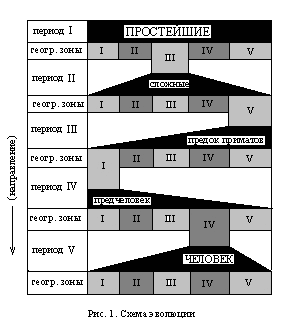
"Представим себе нашу планету упрощенно. Допустим, что есть на ней всего лишь ять географических зон, И пусть на нашей планете появились простейшие... Их появление знаменует собой начало I периода. Отсутствие конкурентной среды (золотой век!) позволило им беспрепятственно освоить все разнообразие имеющихся на планете географических зон (в нашем случае - условные пять). Но разнообразие этих зон детерминировало генетический отбор в соответствующем направлении, что в итоге привело к тому, что простейшие каждой зоны стали приобретать свои отличительные признаки, свойственные только им. Конечным результатом такого процесса было образование первых видов простейших (в нашем случае: условные пять зон - условные пять видов). Один из этих видов в результате такой эволюции приобрел более широкий спектр приспособительных функций, чем другие - превратился из простейшего в нечто более сложное и допустим, что это произошло в III географической зоне. Отметим также и ту очевидность, что филогенетическое преобразование простейших в некое видовое многообразие привело постепенно к тому, что межвидовое размножение стало невозможным, а если даже и да, то этому мешали новоприобретенные физиологические особенности, заставлявший каждый индивид, в силу выживания, гнездиться рядом с таким же, как и он - т. е. колонизировать среду обитания. Далее логика событий рисует нам следующую картину. Сложные, обособившиеся в III зоне, как более сильные и более выживаемые, начали мигрировать в остальные зоны, вытесняя и уничтожая своих собратьев. Вытеснение и уничтожение не приводило к полному истреблению - оно означало лишь то, что хотя остальные виды и продолжали параллельное существование, эволюционно-генетическое преобразование их значительно замедлялось. После распространения сложных по всем пяти географическим зонам (которых в действительности было значительно больше) процесс эволюционно-генетического отбора повторился. Оговоримся, что и этот, II период филогенеза мы значительно упростили, как по горизонтали (количество географических зон), так и по вертикали, ибо он содержит в себе не менее одиннадцати периодов (проторезойский, кембрийский, силурский, девонский, карбонский, пермский, триасский, юрский, меловой, третичный и четвертичный)... Представим себе, что благодаря случайному стечению обстоятельств сильнейшими оказались сложные, живущие в V зоне. Назовем тот вид животных, в которых в этой зоне превратились сложные, предком приматов. И, действительно - пройдя в течение многих миллионов лет через ряд эволюционно-генетических преобразований, они могли в конце третичного периода в какой-либо из географических зон (например, в одном из районов Восточной Африки) превратиться именно в такое животное. И оно, будучи предком приматов, превосходя другие виды в находчивости, ловкости, хитрости и сплоченности, начало экспансивную миграцию из V зоны в остальные и, расселившись, начало эволюционировать в соответствии с вариациями внешней среды. В одной из зон (допустим в I) предок приматов превратился в вид, который мы условно назвали предчеловеком (III период филогенеза). И, наконец, еще одна метаморфоза привела к тому, что предчеловек превратился в человека (IV зона, IV период). В настоящее время (V период) практически нет такого места на земле, где бы не обитал человек. Период этот еще не закончился и он резко отличается от характера эволюционно-генетических процессов прошлых периодов филогенеза. Прежде всего тем, что несмотря на то, что фенотипические формы эволюции успели привести человека к расовым, национальным и прочим видам межзонального различия, генотипические ее формы ведут к увеличению мощи и силы разума, который в противовес агрессивности и антагонистичности не дал и не дает человеку дифференцироваться на несовместимые виды, ибо в любом случае способствует... ...наследственно-генетическому взаимопроникновению". Здесь мы описали феноменологически течение эволюции во времени в соответствии с одной из многих существующих научных гипотез. Надо сказать, что схематически описанный нами процесс вполне укладывается в рамки дарвинизма и сегодня никого удивить не может. И конечно же не ради того, чтобы у кого-нибудь от удивления поднялись брови, мы предпринимаем очередное наше путешествие в незнаемое. И мы, конечно же, не намериваемся опровергать Дарвина. Эволюция есть и этот факт не подлежит никакому сомнению. Однако наряду с неопровержимостью самого этого факта, пугает не столько многообразие гипотез, пытающихся объяснить конкретные пути эволюционного процесса, сколько возможность их легкого опровержения. Ну, например, одна из таких гипотез, утверждает что в результате быстрого погружения дна океана, многие животные оказались вдали от береговой кромки, что изменило само по себе условия их существования настолько, что они мгновенно вымерли и на смену им пришли новые виды, сумевшие приспособиться и выжить. Когда начинаешь интересоваться сколько понадобилось геологического времени для такого быстрого опускания и мгновенного вымирания, то оказывается, что не менее 50000 лет. Теоретически такого временного промежутка вполне достаточно, чтобы мышь в результате эволюционных преобразований могла превратиться (в смысле размеров) в слона, а такое животное, как например, кит - в мышь. Но если это так, то возможно ли объяснять массовую гибель животных (палеонтологически доказанную) и возникновение новых видов изменением условий существования? Строго мыслящие исследователи, принимая во внимание продолжительность катастрофы, выдвигают более подходящую гипотезу, как например, уважаемый нами Д. Соболев. Он утверждает следующее: "Возможность обращения развития обходит значительную часть тех препятствий, какие возникают при отыскании родоначальных форм различных филетических ветвей ныне живущих и ископаемых организмов не среди фантастических и нежизненных схем, а между действительно существовавшими живыми и специализованными формами. Однако, одна эта возможность сама по себе еще не устраняет всех затруднений. Если бы обратное развитие происходило медленно и постепенно, в таком случае мы легко находили бы в ископаемом состоянии следы таковой деградации в виде постепенных переходов между теми группами организмов, которые относятся друг к дружке, как предки к потомкам. В действительности мы этого не наблюдаем. Факты палеонтологии не укладываются в рамки кунтактивной эволюции - прямой или обратной" - [5, с. 133]. Под кунтактивностью здесь понимается постепенность процесса. Конкретизируя свою мысль до наглядного представления того как может осуществляться такое возвращение специализированных организмов к примитивным формам, Соболев предполагает, что в филогении явление обратного развития проявляется в том, "что в филетическом ряду каждый последующий член заканчивает свое онтогенетическое развитие - в отношении всех или только некоторых признаков - на более ранней стадии, чем предшествующий член: в последовательном ряду поколений потомки "неотенически" не дорастают до тех стадий, на которых стояли предки" - [5, с. 104, 105]. Под неотенией в данном случае разумеется возможность более раннего развития и полового созревания. С таким предположением ученого, несмотря на то, что он приводит много палеонтологических примеров, можно и не соглашаться, исходя из доводов пригодных для абсолютно противоположной точки зрения, но и плюс к этому достаточных для защиты иных гипотез, имеющих такое же право на существование, как и представленная Соболевым. Если отбросить его гипотезу, как неподтвержденную более убедительными фактами, чем теми, которые подчеркиваются в его работе (например, "что птеридоспермы (Cycadofilices) появились задолго до возникновения настоящих папоротников, так что высший тип исторически предшествует низшему" - [5, с. 110], то что остается? Наличие существенных результатов наблюдений над современными популяциями, экстраполяция которых на более длительные периоды времени убеждает в том, что эволюция на протяжении миллионов лет скорее всего происходила путем постепенных прогрессивных изменений. Однако данные палеонтологической летописи, особенно по морским беспозвоночным, свидетельствуют о параллельных возможностях совершенно иных процессов. Элдредж и Гулд (Eldredge, Gould, 1972) отметили, что среди ископаемых форм, лишь очень немного надежно документированных случаев существенных изменений вида с течением времени. Наиболее обычные из них сохраняют свои характерные морфологические особенности на протяжении миллионов лет. Когда же изменения все-таки наступают, они, насколько можно судить по находкам, требуют относительно короткого периода по сравнению с общей продолжительностью существования вида. И это практически то единственное, что нам достоверно известно. Не потому ли, например, располагая практически одной и той же информацией, одни ученые приходили к выводу, что внутри вида Homo erectus объем черепной коробки существенным образом не менялся (Rightmire, 1981), тогда как другие (Cronin et al., 1981) настаивали на постепенных изменениях в ряду от Australopithecus до Homo sapiens? Исходя из отмеченного нами спорного опыта предшественников, сразу же оговоримся, что, внося свой скромный и, весьма вероятно, далеко не академический вклад в гипотетические построения апостолов научной мысли, мы не собираемся опровергать ни одной из существующих гипотез. Очень возможно, что все они (не станем утомлять читателя их перечислением) имеют вполне законное право на существование - хотя бы по той явной причине, что эволюция в способах восхождения от низшего к высшему не имеет ограничений в выборе путей развития. Подобная недетерминированность характерна, прежде всего, для позвоночных. И связано это очевидно с геометрической особенностью первого деления оплодотворенного яйца. Если у специализированных форм различных групп беспозвоночных, включая кольцевых червей, членистоногих, и головоногих моллюсков первые деления яйца происходят так, что клетки (бластомеры) следующего по счету ряда вклиниваются между клетками предыдущего (это называется спиральным дроблением) и, таким образом, судьба каждой клетки предопределена с самого начала, т. е. если один из бластомеров удалить, соответствующая ему часть организма попросту не разовьется; то у хордовых и иглокожих, родоначальников всех существовавших и существующих ныне позвоночных, "первые клеточные деления протекают по типу радиального дробления, т. е. бластомеры располагаются точно друг над другом. Если одну из клеток на ранних стадиях онтогенеза удалить, соседние могут компенсировать ее недостачу и разовьется нормальный зародыш (недетерминированное деление)" - [4, т. I, с. 30]. Нам кажется, что выход хордовых и иглокожих в эволюционном процессе на недетерминированное деление клеток стал возможным только потому, что биохимическое образование правых и левых молекул в живых организмах характеризуется несимметричностью (впервые замечено Пастером), которая сдвигает любую живую систему в сторону неустойчивости и посему делает ее максимально способной к новым формообразованиям в соответствии с изменениями внешней среды. Уделяя чрезвычайное внимание этой несимметричности, В.И. Вернадский отмечает в связи с этим неравенство правых и левых спиралей в форме организмов, выявляемое "в редком проявлении "левшей", причем для некоторых организмов левши являются преобладающими и могут быть принимаемы как видовой признак" - [2, кн. 1, с. 58]. Здесь следует отметить замеченную нами идентичную несимметричность в мире насекомых и в мире самых высокоразвитых животных, проявляемую в расположении органов воспроизводства звука. У кузнечиков, например, правое надкрылье находится под левым. Указуя на эту природную закономерность, Фабр Ж. А. В своей знаменитой книге "Нравы насекомых" пишет - "смычок левого надкрылья есть двигатель; терка правого надкрылья - точка колебания; натянутая кожица зеркальца - резонатор, вибрирующий при посредстве своей приводимой в сотрясение рамки" - [6, т.II, с. 388]. Источник порождения звуковых сигналов несимметричен и у китов. "Направленность посылаемого сигнала контролируется специальным образованием, расположенным на голове около ноздри. У современных одонтоцетов это одно отверстие ( у остальных китов их два), и его модификация приводит к заметной асимметрии окружающих костей. Такая асимметрия возникла независимо шесть, а то и десять раз при формировании различных семейств одонтоцетов. Можно сгруппировать зубатых китов в пять надсемейств, соответствующих крупным филогенетическим радиациям. Каждое надсемейство характеризуется особым типом асимметрии" - [4, т. III, с. 94]. После наших искренних уверений, что мы не собираемся опровергать ни одной из имеющихся гипотез в области эволюционного изменения видов зачем, собственно говоря, нами затеян весь этот разговор? Конечно же не для того, чтобы удивить читателя эрудицией, ибо, очевидно, что какой бы обширной она не казалась, она черпает информацию из готовых источников знаний, а наша задача внести в эту копилку нечто новое. Исследуя ту проблему, в которую мы, со всей очевидностью, уже углубились и проанализировав сложившуюся ситуацию, мы можем с уверенностью сказать, что все существующие гипотезы выражают только одну сторону какого либо палеонтологически фиксируемого явления и посему одна гипотеза не походит на другую и ей противоречит. Исходя из этого положения, бросающегося в глаза, назрело, вероятно, время для такой теоретической основы, которая бы тяготела не к индивидуальной оригинальности, но стремилась бы удовлетворить все прежние гипотезы - т. е. не являлась бы по отношению к ним противоречивой. Для того, чтобы приблизиться к поставленной цели необходимо определить недостаток всех предшествующих теорий и недостаток этот должен быть в равной мере присущ без исключения им всем. Нам кажется, что все существующие гипотезы, относящиеся к путям и возможностям эволюционных преобразований, страдают односторонним подходом к проблеме, заключающемся в том, что рассматривается сам факт морфологического изменения и предполагаемое соответствующее влияние внешней среды, и, исходя из этого плоскостного подхода фантазируется та или иная возможность, когда сущность явления заключается в том, чтобы ответить на коренной вопрос - что происходит с изменяющимся организмом внутри, каков порядок эволюционного преобразования - изменение внешнего воздействия (условий среды обитания) и в результате последующее приспособительное изменение организма, или, как мы предполагаем, существует третий фактор - промежуточный и самый важный - процесс реагирования организма, который необходимо рассматривать в самой его материальной сущности? |
 |
 |
 |
 |