|
 |
 |
 |
Александр Вихров Где искать следы иного разума?Более 40 лет уже ученые ведут активные поиски внеземной жизни. В их распоряжении мощнейшие радиотелескопы, периодически посылающие послания в различные области галактик, где предполагается существование разумных цивилизаций. Публике хорошо известен глобальный научный проект SETI по поиску сигналов иного разума. Миллионы долларов уже потрачено и еще много запланировано потратить. Только кажется, что эти деньги тратятся впустую. Если внеземной разум и существует, то ему гораздо легче найти нас, чем нам его. Вполне возможно, что земля уже входит в сферу его деятельности. Многие наверное помнят нашумевший когда-то фильм Дэникена "Воспоминания о будущем". Публике были предъявлены, так сказать, вещественные доказательства пребывания на Земле в далеком прошлом представителей развитых инопланетных цивилизаций. Многие из этих вещдоков были сразу же опровергнуты, другие вызвали немало споров. Можно по разному трактовать рисунки инков и происхождение "взлетных полос" плоскогорья Наска. Для кого-то они выглядят убедительным подтверждением инопланетного присутствия, для других нет. Частые посещения Земли неопознанными летающими объектами также будоражат воображение многих ее обитателей. К тому же НЛО порой оставляют следы - правильные круги на полях, оплавленные породы с наведенной радиоактивностью и, как утверждается, искореженные в результате катастроф кусочки собственных конструкционных материалов. Итак, искомые "следы" вроде имеются. Но как к ним относиться? Много всякой чепухи здесь намешано. Каким образом можно отделить зерна от плевел? Ниже я попробовал сформулировать критерии, по которым можно судить о принадлежности тех или иных объектов к предметам искусственного внеземного происхождения:
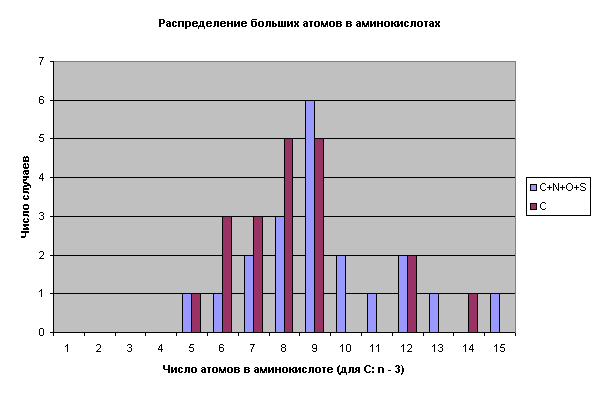
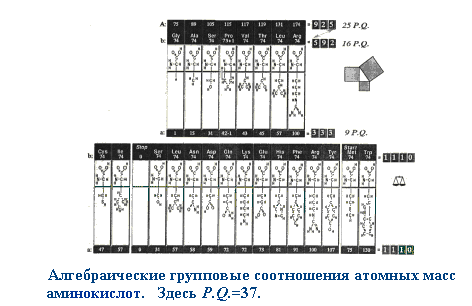
Полагаю, что представленные критерии достаточно жесткие. Давайте попробуем оценить по ним объекты, претендующие на инопланетное происхождение. Пирамиды Хеопса тянут максимум на 6-7 баллов за счет своей явной искусственности. "Космодром" Наска - на 5, остатки загадочных материалов с места "катастрофы" НЛО в районе Дальнегорска - всего на 4. Не густо. Есть ли на Земле вообще объекты, удовлетворяющие всем 9 критериям? Да есть. Это белки и нуклеиновые кислоты, из которых состоят живые организмы. Разумеется, соответствие всем вышеописанным критериям не является доказательством их искусственного происхождения. Оно лишь подсказывает, в каком направлении следует искать. Давайте попробуем поискать другие свидетельства. Итак, живые организмы состоят из тех же атомов, что и неживые, но совершенно других молекул. Кардинальное отличие - односторонняя симметрия структурных элементов белков и нуклеиновых кислот, огромная молекулярная масса биополимеров. Что представляет собой белковая молекула? Ниточный полимер из разнородных нерегулярных звеньев - аминокислотных остатков. Аналогов подобному в природе нет. Удивительно, что эта нерегулярная аминокислотная последовательность соединяется исключительно регулярной пептидной связью - СO - NH - , все атомы которой лежат в одной плоскости. Образование пептидных цепочек аминокислот в водных растворах противоестественно ( вопреки принципу Ле-Шателье), намного предпочтительнее возникновение водородных и дисульфидных связей между атомами боковых ветвей. Налицо неестественная и избирательная упорядоченность атомов в белковой молекуле. Метод формирования белков не имеет аналогов в природе. Углерод в органических соединениях образует 4 ковалентные связи и в соответствии с принципами кристаллографии, его полимер должен расти во всех четырех направлениях, формируя древовидные и сеточные структуры. Ни один кристалл не растет таким образом, чтобы подобно протеинам, сначала формировалась длиннющая нить толщиной до одного атома, а затем она складывалась, образуя плотное тело. Формирование белковой глобулы скорее напоминает вязание свитера, чем рост кристалла. Обратимся к аминокислотному набору - базовым элементам белковой структуры. Обращает на себя внимание их дифференцированный выбор, предполагающий многоступенчатый отбор элементов из неограниченого исходного числа компонентов. Во-первых, это собственно аминокислоты, аминокислотный ряд, не включающий другие органические кислоты, нуклеотиды и прочее. Во-вторых, только 20 наименований из сотен возможных и существующих в природе. В-третьих, только α- модификации ( искючаются β, γ и другие изомеры). В-четвертых, только молекулы, имеющие пространственную L- ориентацию (D-ориентация абсолютно исключена). Каковы возможные механизмы подобного отбора? В природе наблюдается отбор по подобию (идентичности), совпадению химико-физических свойств, размерам и сходных условий формирования. Давайте проанализируем принципы отбора аминокислот стандартного ряда. Белковые аминокислоты легко поддаются классификации по структуре и свойствам их боковых цепей. Есть несколько методов классификации, наиболее показательным я считаю следующий. Аминокислотный набор распределяется на следующие группы: алифатическую (чисто углеводородная боковая цепь), содержащую 6 аминокислот, углеродзамещенную (боковая цепь содержит другие атомы, замещающие в ней углерод) - 10 аминокислот, и ароматическую - 4 аминокислоты. Первая пара алифатических аминокислот - самые простые - глицин и аланин, не имеющие собственных изомеров (аланин - ввиду L-ориентации). Следующий по сложности довольно простой гомолог, содержащий углеводородную цепочку из двух атомов углерода почему-то отсутствует, зато потом следует по два С5 и С6 изомера. Боковая цепочка одного из С5 изомеров замыкается на атом азота, образуя циклическую группировку (иминокислоту пролин). В углеродзамещенной группе четко прослеживается парность аминокислот: 2 серосодержащие аминокислоты, 2 - содержащие гидроксильную группировку OH, по паре аминокислот с выраженными кислотными и основными свойствами, 2 аминокислоты содержат амидные группировки. Принцип отбора - каждой твари по паре. Обращает на себя внимание большое разнообразие концевых группировок аминокислот, что обеспечивает богатое разнообразие свойств. Оставшаяся небольшая ароматическая группа аминокислот не сохраняет принцип парности, зато остается принцип разнообразия: простое бензольное кольцо, затем добавляется основная азотсодержащая группировка, следующая аминокислота содержит бензольное кольцо с гидроксильной группой и наконец - двойное ароматическое кольцо. Это разнообразие еще более удивительно, если учесть явную ограниченность и консервативность химического состава самих аминокислот. Их состав можно выразить формулой 4 + 1, где 4 - количество обязательных атомов = числу атомов пептидного звена (- СO - NH -) + 1 дополнительный. Эта закономерность справедлива и для нуклеотидов - там дополнительным элементом является фосфор, а не сера, как у аминокислот. Характерно, что и число стандартных нуклеотидов также укладывается в эту формулу: исключительно по 4 - во всех нуклеиновых кислотах, но в РНК один из них (тимин) заменен урацилом. Поэтому, эту зависимость можно считать константой наряду с константами, выражающими число нуклеотидов в НК и аминокислот в белках. Все три константы связаны арифметически: 4 х (4+1) = 20. На рис. 1 показано распределение аминокислот стандартного набора по числу содержащихся в них атомов углерода, равно как и суммы "больших" атомов (C, N, O, S).
Рис. 1. Хорошо заметно неравномерное распределение аминокислот по углероду, которое далеко от естественного. На первую половину диаграммы (3-6 атомов С) приходится 17 аминокислот и только 3 - на вторую (7-9 атомов С). Непонятен провал гомологического ряда между С6 и С9. Этот провал можно объяснить усиливающейся тенденцией к замене атомов углерода другими его аналогами. Это и обусловливает широкое разнообразие свойств боковых цепей аминокислот. Анализ аминокислот стандартного ряда показывает, что отмеченные ранее природные принципы отбора не могут объяснить их уникального разнообразия. Действительно, стандартные аминокислоты сильно различаются по строению, размерам, свойствам и сложности их образования. Реальным принципом отбора аминокислот является обеспечение максимально возможного разнообразия химических свойств концевых группировок боковых цепей. Парный характер подбора это разнообразие закрепляет, стабилизирует. Достаточно ли этого для утверждения искусственности белков и НК? Думаю, что нет. Поищем другие закономерности. |
 |
 |
 |
 |